
The molecular basis of force selectivity by PIEZO2
Article Date: 04 March 2026
Article URL: https://www.nature.com/articles/s41586-026-10182-7
Article Image: Figure 1
{kind=link}
Summary
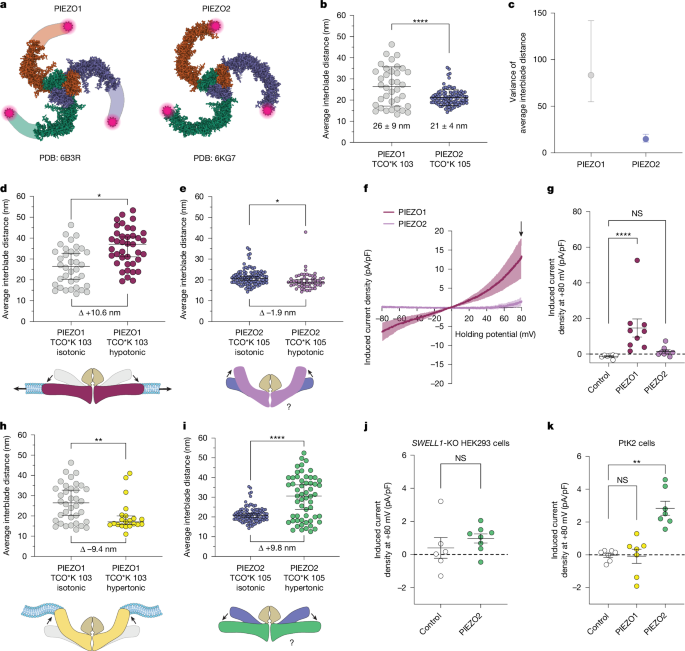
This Nature paper dissects why PIEZO2, a key mechanotransduction channel in touch and proprioception, is tuned to indentation rather than to uniform membrane tension like PIEZO1. Using cutting-edge MINFLUX nanoscopy combined with fluorogenic DNA‑PAINT, the authors measured single‑molecule conformations of PIEZO1 and PIEZO2 in cell membranes. They show that PIEZO2 is mechanically tethered to the cortical actin cytoskeleton through the scaffold FLNB. This tethering (via the intracellular IDR5 region and FLNB) immobilises PIEZO2, biases its blades into a pre‑expanded state and thus lowers the threshold for indentation. Disrupting actin, deleting IDR5 or removing FLNB makes PIEZO2 behave more like PIEZO1: blades expand with membrane stretch and the channel becomes sensitive to bilayer tension. The study combines structural nanoscopy, electrophysiology in SWELL1‑KO and FLNB‑KO cells, cross‑linking mass spectrometry, siRNA/CRISPR perturbations and in situ co‑localisation in mechanoreceptor endings to build a tether‑coupled gating model for PIEZO2.
Key Points
- PIEZO2 differs from PIEZO1: PIEZO1 is gated primarily by lateral membrane tension; PIEZO2 is preferentially gated by local indentation forces.
- High‑precision MINFLUX + fluorogenic DNA‑PAINT measured interblade distances and mobility of individual channels in intact cell membranes.
- PIEZO2 blades are more conformationally rigid and less expanded at rest than PIEZO1; FLNB tethering biases PIEZO2 blades into a pre‑expanded, sensitive state.
- FLNB was identified as a molecular tether by cross‑linking MS and functional siRNA/CRISPR screens; FLNB loss increases PIEZO2 mobility and renders it stretch‑sensitive.
- Disruption of actin or deletion of PIEZO2 IDR5 converts PIEZO2 to a force‑from‑lipid behaviour, increasing stretch sensitivity and mobility.
- PIEZO2 and FLNB co‑localise in peripheral mechanoreceptor terminals (Meissner corpuscles and lanceolate endings); FLNB knockdown alters endogenous neuronal PIEZO2 force selectivity.
- Overall model: PIEZO2 senses membrane deformation relative to an actin tether (force‑from‑filament/tethered gating), explaining its heightened sensitivity to poking/indentation.
Content summary
The authors used PtK2 cells for near‑planar membrane imaging and genetically encoded click‑labelling at equivalent extracellular blade loops of PIEZO1 and PIEZO2. Fluorogenic DNA‑PAINT increased photon yields and resolution when used with MINFLUX, enabling reliable 3D localisation of blade positions and single‑molecule tracking in live cells.
Key experimental observations:
- Without stimulation, PIEZO2 blades are significantly less expanded and explore a narrower distribution of conformations than PIEZO1, indicating higher rigidity.
- Hypo‑osmotic swelling (which increases membrane tension) expands PIEZO1 blades and opens the channel, but compacts PIEZO2 blades and does not activate it—opposite responses to the same stimulus.
- Hypertonic shrinkage expands PIEZO2 blades (contrary to expectation) and can activate PIEZO2 in cells with a stiff cortex, implicating cytoskeletal coupling in force transmission.
Functional tests revealed that disrupting F‑actin (cytochalasin D or latrunculin A), deleting IDR5 or knocking down/out FLNB increases PIEZO2 blade expansion with membrane stretch, increases channel mobility and enables stretch‑evoked currents. Cross‑linking MS followed by knockdown screens singled out FLNB as the tethering partner; FLNB‑KO or knockdown reproduces the actin‑disruption phenotype. smFISH and STED imaging show PIEZO2 and FLNB co‑expression and nanoscale co‑localisation in low‑threshold mechanoreceptors, and FLNB knockdown in DRG neurons makes endogenous PIEZO2 responsive to membrane stretch.
Context and relevance
This work resolves a long‑standing paradox: PIEZO2 is exquisitely sensitive to poking/indentation in sensory neurons yet appears relatively insensitive to bilayer tension in patch assays. The study provides a mechanistic explanation—an actin‑FLNB tether changes how mechanical energy is delivered to the channel, prioritising indentation over uniform tension.
Relevance to the field:
- Explains modality specificity in mechanosensation (how similar channels can detect different mechanical cues).
- Suggests a general organisational principle: pairing PIEZOs with local ultrastructural partners (filamins, tethers or extracellular anchors) tunes mechanical sensitivity and threshold.
- Has implications for sensory physiology and disease: FLNB and PIEZO2 mutations produce overlapping phenotypes, hinting at shared pathways in touch, proprioception and respiratory control.
Author’s take
Punchy: this is a clean, elegant piece of work. The authors marry single‑molecule nanoscopy and solid electrophysiology to move beyond static cryo‑EM snapshots and show how cellular context — a simple actin scaffold — rewires channel mechanics. If you work on mechanotransduction, sensory biology or ion‑channel gating, the details here are highly actionable: FLNB and IDR5 emerge as real levers for tuning PIEZO2.
Why should I read this?
Because it explains why PIEZO2 behaves weirdly in different assays — and it does so with single‑molecule precision. It’s the kind of paper that changes how you design experiments: want to study touch sensitivity or tweak channel thresholds? Consider the cytoskeletal scaffold. Also, the techniques (MINFLUX + fluorogenic DNA‑PAINT) are a neat template if you’re after nanometre‑scale conformational readouts in cells.